Chapter 11: Selective Distribution of Ions, Sugars, and Free Amino Acids
Cardinal adsorbents "expose" solute adsorption sites.

As always, this is not medical advice, and reading this does not form a client relationship with me - your health is your responsibility.
I am decreasing consults as we get closer to September. However, I started a Discord server. If you have health and wellness questions, etc., please consider joining - this would be the best way to support my work! Many have asked for my “thoughts” on things. The best place to start would be some of my older Substacks - especially the ones on supplementation (1 and 2). Also, please check out my IG story highlights.
Today’s Substack will continue with Chapter 11, Selective Distribution of Ions, Sugars, and Free Amino Acids. Chapter 10 discussed a different view of ATP as the driver of biological work performance.
Summary:
Chapter 11 contains more mathematical theory and plots depicting data sets. Adsorption of intracellular solutes, like D-glucose, K+, etc., are made more likely under cardinal adsorbents like insulin and ATP. “This chapter dealt with the equilibrium (or steady-state) distribution of solutes. The next chapter uses the same basic concepts to describe the kinetic properties of solutes (and solvent) movements into and out of cells (i.e., their permeabilities).” The figures I shared from the chapter are the most ellucidating and help to summarize the equilibrium state.
11.1 The General Theory of Solute Distribution
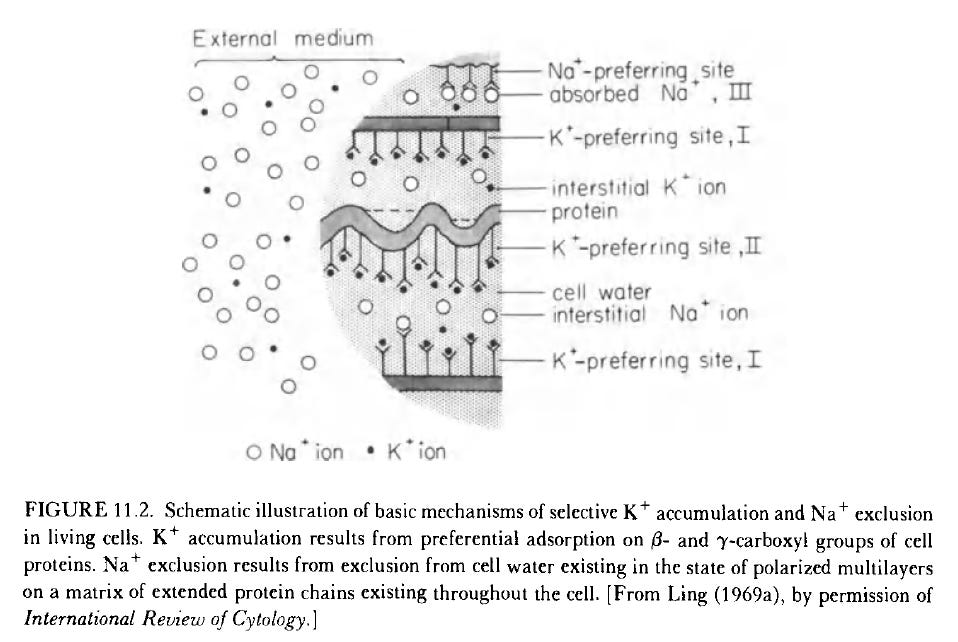
Even though there is a greater concentration of Na+:K+ external to the cell, there is a greater K+:Na+ inside the cell. The K+ is mostly adsorbed to the cell’s amino acids’ beta- and carboxyl-groups. The free Na+ and K+ in the cell have to do with their external concentration.

Shown above: The cardinal adsorbent, ATP, change the folding of the amino, which causes the cellular water to polarize and form structured multilayers, and adsorbed ions, like K+. For the solute sites (e.g., K+) to adsorb, the cardinal adsorbent must occupy the cardinal site. “In dead cells, water is no longer polarized in multilayers because the cardinal adsorbent essential for the maintenance of the proper extended conformation of these matrix proteins is depleted.” This is why I focus on the TCA and ETC as they are known to lead to the production of ATP - ATP, amino acids, and electrolytes lead to biological work performance, the high-energy resting state. The phosphocreatine system, etc., is also part of ATP regeneration. And there are other processes like glycolysis and fatty acid oxidation that feed into TCA and ETC.
11.2 Experimental Testing of the Theory
The order of cell solute adsorption is Rb+ > K+ > Cs+ > Na+ in a “structured cell.” As cells gain Mg+2 (K+), the intracellular K+ (Mg+2) concentration remains unchanged. This is because amino acid beta- and gamma-carboxyl groups adsorb K+ “ and pairs or groups of chelating anionic sites for Mg+2.” Anionic means negatively charged. When Mg+2 is adsorbed intracellularly, an equal amount of Cl- or SO₄²⁻ is also. SO₄²⁻ is the sulfate ion. These enter without exchanging other ions - where are all the ion swaps from the “pumps?”
Damaging a cell causes a local phase change causing “the physical state of water from its normal polarized multilayered state to one closer to normal liquid water” occurs.
Intracellular K+ accumulation is akin to oxygen binding to hemoglobin.
The living cell is a “protein-ion-water system [that] is a three-dimensional cooperative assembly and, as such can undergo a sharp transition between two alternative states with a relatively minor temperature changes.” The lower the temperature, the less the intracellular K+ and the more Na+ comes into the cell. This is especially seen at ~13C (55F), but shifts start around body temperature, 37C (98.6F) - a change towards more intracellular Na+:K+. Models using water polarized by polymers “remains unchanged from 0C to much higher temperature. However, at 35C, the polymer-water system undergoes an abrupt change. As a result, the bulk of the water becomes totally normal; indeed, it separates from the polymer-water that now exists as a separate coacervate phase.” This is why the acceptance of dropping body temperatures is a move in the wrong direction. In frog sartorius muscles, there is an “abrupt increase of sucrose concentration in the muscle at temperatures above 37C… at 40C [104F] the sucrose concentration equals that of the external medium.” Sucrose is glucose and fructose. This should give someone pause when they have a fever. Ling shows that cells do not require insulin for sucrose to enter. We now look to glucose transporters to explain why - GLUT4 is insulin-dependent. However, Ling argues, “the low concentration of sucrose in normal cells is due to the solute exclusion property of polarized water; this property is altered at high temperature in an all-or-none manner.” The thyroid is like the gas pedal for glucose metabolism and is a proxy for body temperature. When glucose metabolism is not “properly” occurring, this will decrease thyroid and body temperature. People misunderstand this order of operations and aim to correct the issue by giving thyroid when this is the incorrect starting point. This alone would explain why many do not do well when FT4, FT3, and rT3 levels are “optimized” and why many do not do well with thyroid medication - yes, even T3.
Cardiac glycosides like strophanthus (ouabain seems the causative agent - Ling refers to it as another cardinal adsorbent) increase and decrease intracellular Na+ and K+, respectively. It does not affect ATP concentration until high doses. Increased intracellular Na+ typically goes with increased intracellular Ca+2. This is seen with cardiac glycosides and leads to muscle cells having a faster and larger contraction at a decreased heart rate = higher cardiac output with no increase in blood pressure.
In vitro experiments show increasing the extracellular [K+] or [Ca+2] retains intracellular K+. Intracellular K+ is conserved by extracellular Ca+2 > Sr+2 > Co(NH3)5Cl+2 > Mg+2 > 0.
“The extremely high binding constant of ATP on myosin also leaves no room for existence of a substantial concentration of free ATP in cells; virtually all ATP must also be adsorbed.” “As ATP declines, there is also a progressive depolarization of water and a progressive rise” cells uptaking Na+ and sucrose. Cyanide blocks the regeneration of ATP, leading to a rapid increase in free intracellular Ca+2.
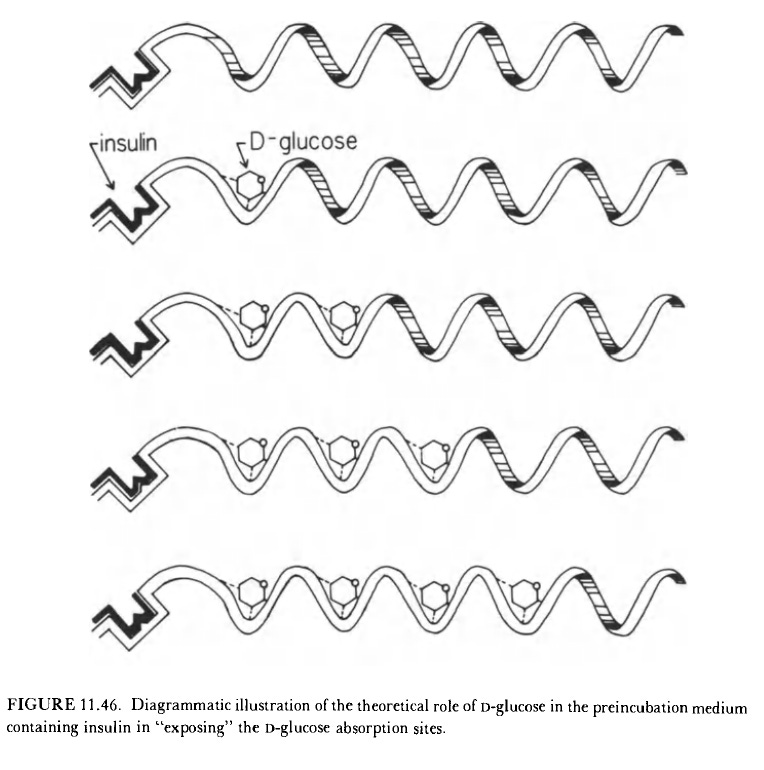
Ling considered insulin to be another cardinal adsorbent like Ca+2 and ATP. “Cardinal sites that adsorb and interact with drugs and hormones are obviously identical with what have long been capped receptor sites by pharmacologists. Yet in terms of the AI hypothesis, cardinal sites reacting with ATP, Ca+2, drugs, and hormones are basically alike.” “Part of the glucose is dissolved in cell water (which under normal conditions remains relatively constant), and part is adsorbed onto protein sites (which are under the control of cardinal sites interacting specifically with insulin)” - “the experimental studies of D-glucose accumulation in the cell are entirely analogous to those of K+ and Na+ and are not “membrane phenomena”.” Cells need to be exposed to enough D-glucose for it to be taken up - Randle cycle idea.